종양이 자신의 성장을 유지하기 위해 혈관이 필요하다면, 어떻게 그 필요성을 달성할 수 있을까요? 첫 번째 힌트는 세포가 통과할 수 없을 정도의 작은 구멍을 가진 필터로 둘러싸인 미세용기의 내부에 암세포를 놓아두었던 연구에서 도출되었습니다. 이런 용기를 동물의 체내에 이식하면, 주위의 숙주조직에서 새로운 모세혈관들이 분열하기 시작합니다. 이와는 대조적으로, 정상 세포를 담은 용기를 같은 방식으로 이식했을 경우에는 혈관의 성장이 촉진되지 않았습니다. 이는 암세포가 만들어내는 분자가 필터의 작은 구멍을 통해 확산하여 주변의 숙주조직의 혈관신생을 활성화한다고 설명할 수 있습니다.

다음 연구는 혈관신생을 촉진하는 데 관여하는 분자를 규명하는 것으로 많은 연구자들이 수십 년간 이 연구에 전념하고 있습니다. 결국 이런 집중적인 노력으로 혈관신생을 활성화하는 몇몇 작은 분자들과 12개 이상의 단백질이 규명되었습니다. 이들 단백질 중 2개의 단백질은 혈관 내피세포 성장인자와 섬유모세포 성장인자로서 종양성장을 지속하는 데 특히 중요한 것으로 여겨지고 있습니다. 혈관 내피세포 성장인자와 섬유모세포 성장인자는 많은 종류의 암세포에서 합성되며 이들은 내피세포의 표면에 위치한 특정 수용체 단백질에 결합하여 혈관신생을 유도합니다.
이런 과정이 어떻게 일어나는지 보기 위해 잠시 VEFG에 집중해봅시다. VEGF는 대부분의 종양에서 생성되고 주변조직으로 분비됩니다. VEFG분자가 내피세포와 만나면, 내피세포의 표면에 있는 VEFG 수용체에 결합하여 이를 활서오하시킵니다. 활성화된 수용체로부터 일련의 경로를 통해 신호전달단백질로 전달된 신호는 그 후 세포의 행동과 유전자의 발현 변화를 유도합니다. 그 결과 내피세포는 분열을 시작하고, 주변 조직으로 분비되면서, 단백질을 분해시키는 일련의 효소인 matrix metallo-proteinase(MMP)군을 합성합니다. MMP들이 이웃 세포 사이의 공간을 채우고 있는 세포외 기질의 구성 성분을 분해하므로, 내피세포가 주변조직으로 이동할 수 있게 됩니다. 이때 증식된 내피세포는 빈 혈관으로 들어가 새로운 혈관 망상조직으로 발달합니다.
많은 종양에서 VEGF나 FGF를 합성한다 할지라도 이것이 혈관 신생 활성에 대한 유일한 설명이 되지는 못합니다. 혈관신생이 진행되기 위해서는, 이들 분자가 정상 상태에서 혈관의 성장을 억제하는 혈관신생 억제물질들의 효과를 극복해야만 합니다. 혈관신생을 자연적으로 억제하는 안지오스타틴, 엔도스타틴, 트롬보스폰딘을 포함하는 12개 이상의 혈관억제물질들이 밝혀졌습니다. 혈관신생의 억제물질들과 활성인자의 농도가 정교하게 균형을 이루어, 종양의 새로운 혈관 성장이 유도될 것인지의 여부가 결정됩니다. 종양이 혈관신생을 자극할 때에는 혈관신생 활성인자의 생성량도 증가하며, 그와 동시에 혈관신생 억제물질의 합성은 저하됩니다.
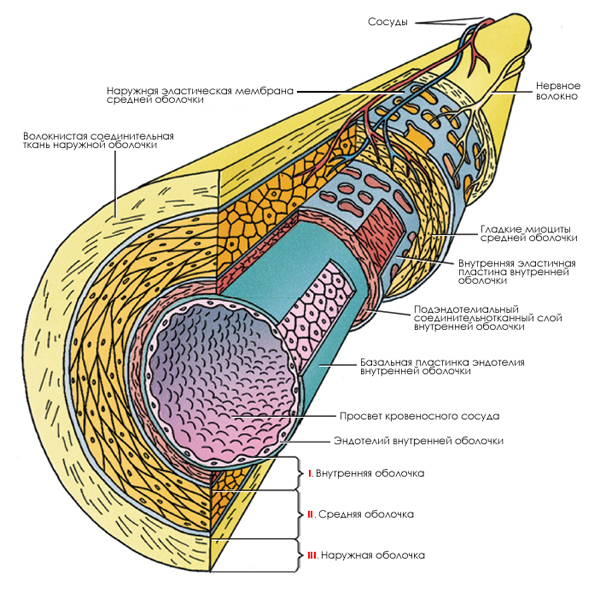
종양 혈관 신생.
자체적인 혈액 공급이 없을 경우, 종양 덩어리는 일반적으로 1~2mm 지름을 넘지 못하고 작은 크기로 본래 생긴 자리에 국소적으로 머물러 있습니다.
암세포가 혈관신생-촉진 분자들을 합성한다는 증거.
암세포가 투과할 수 없는 작은 구멍을 가진 필터로 둘러싸인 미세용기에 암세포를 두었습니다. 이 미세용기를 동물에게 이식한 경우, 새로운 혈관들이 주변 숙주조직에서 분열하기 시작했습니다. 이러한 관찰을 암세포가 합성한 분자들이 필터를 투과하여 주변조직의 혈관신생을 자금함을 제시합니다.
VEGF에 의해 유도되는 혈관신생의 주요 단계.
암세포는 혈관 내피세포의 표면에 존재하는 수용체 단백질에 결합할 수 있는 VEGF 분자들을 분비합니다. 결합된 VEGF는 그 수용체를 활성화시켜 유전자 발현과 세포행위에 변화를 일으키는 신호전달 단백질의 활성을 이끌어냅니다. 결과적으로 내피세포의 분열과 이동이 촉진되고, 세포외 기질의 구성요소를 분해하는 MMP가 분비됩니다. 분열하는 내피 세포들이 점차적으로 새로운 혈관의 네트워크를 형성합니다.